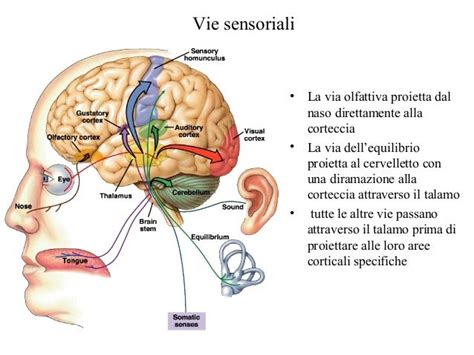
Il senso del tatto, pur apparendo immediato e semplice, è in realtà un sistema straordinariamente complesso, distribuito su tutta la superficie del nostro corpo. La sua funzione primaria è quella di trasformare gli stimoli meccanici esterni in segnali nervosi comprensibili dal nostro cervello, permettendoci di interagire con il mondo circostante e di percepire la nostra stessa posizione al suo interno. Questa complessa rete di sensori, i recettori tattili cutanei, lavora incessantemente per fornire al sistema nervoso informazioni vitali sulla natura e sull'intensità degli stimoli che incontriamo.

La Trasduzione dello Stimolo Meccanico: Dalla Cute al Cervello
Alla base della sensazione tattile vi sono i meccanocettori, recettori periferici specializzati che svolgono il ruolo cruciale di trasduttori. Essi convertono gli stimoli meccanici applicati alla cute, come una pressione, un tocco leggero o una vibrazione, in impulsi nervosi. Questi impulsi vengono poi trasmessi lungo le fibre nervose sensitive fino ai centri nervosi superiori, principalmente la corteccia cerebrale, dove vengono decodificati e interpretati come sensazioni tattili.
L'intensità della sensazione percepita è direttamente correlata alla forza dello stimolo applicato. Tuttavia, i meccanismi precisi attraverso cui il sistema nervoso aumenta la percezione all'aumentare dell'intensità dello stimolo sono ancora oggetto di studio. La capacità discriminativa, ovvero la finezza con cui riusciamo a distinguere stimoli vicini tra loro, è strettamente legata alla densità di recettori presenti in una determinata area cutanea. Le aree con un maggior numero di recettori per unità di superficie, come il palmo della mano, presentano una maggiore capacità discriminativa. È interessante notare come la sensibilità tattile della mano sia ulteriormente potenziata quando il soggetto muove attivamente l'arto, suggerendo un'interazione dinamica tra movimento e percezione.
Il Ruolo della Propriocezione nella Percezione Spaziale
Parallelamente alla sensibilità tattile, un ruolo fondamentale nella nostra percezione del mondo e di noi stessi è svolto dalla propriocezione. Questo senso interno ci permette di percepire la posizione del nostro corpo e delle sue singole parti nello spazio, nonché la contrazione muscolare e il movimento, anche in assenza di input visivi. La propriocezione nasce dalla stimolazione di specifici recettori sensoriali distribuiti in vari organi, tra cui la fascia connettivale, le articolazioni, i tendini, i muscoli, le capsule articolari e la cute stessa.
I recettori coinvolti possono essere classificati in diverse categorie: esterocettori (sensibili a stimoli esterni), propriocettori (situati nei muscoli, tendini e articolazioni) ed enterocettori (situati negli organi interni). Anche i recettori vestibolari dell'orecchio interno, fondamentali per l'equilibrio, contribuiscono a questa complessa rete di informazioni.
La conoscenza della nostra "conformazione" e posizione spaziale dipende da questi recettori periferici. La risposta alla domanda "chi sono io?" è intrinsecamente legata alla risposta alla domanda "dove sono io?". Le rappresentazioni mentali (engrammi), da cui scaturiscono le abilità motorie complesse, nascono infatti dai riflessi estero-propriocettivi.
Dallo Stimolo all'Azione: L'Origine Motoria della Mente
Un aspetto cruciale, spesso sottovalutato, è il ruolo centrale del movimento e dell'azione nei processi di rappresentazione mentale, fin dalle prime fasi dello sviluppo embrionale. L'embrione è, prima di tutto, un organismo motorio. Durante la fase embrionale, fetale e la prima infanzia, l'azione precede la sensazione: movimenti riflessi vengono compiuti e solo successivamente se ne ha una percezione a livello cerebrale. Le funzioni motorie e il corpo, lungi dall'essere entità subordinate alle attività cognitive, ne costituiscono invece l'origine, influenzando persino la formazione del linguaggio, della mente e del pensiero.
La propriocezione può quindi essere definita come la "coscienza di sé". La perdita del controllo sul proprio corpo implica, di conseguenza, una perdita di controllo sui propri pensieri ed emozioni. Come affermato da R. Paparella Teccia nel 1988, "Muscoli, tendini, capsule, legamenti sono organi di senso e organi di moto, impegnati nella ricezione della stimolazione ambientale prima che nell'effettuazione della risposta di moto…il senso non può giammai dissociarsi dal moto".
L'importanza del sistema di feed-back sensoriale è evidenziata dalla predominanza delle fibre nervose sensitive rispetto a quelle motorie in molti nervi. Ad esempio, nel nervo tibiale, la proporzione tra fibre sensitive e motorie è di circa 3 a 1. Ulteriori ricerche sulla corteccia motoria hanno rivelato che essa è organizzata non tanto per aree topografiche corporee, quanto piuttosto in relazione a specifici movimenti corporei complessi, orientati verso un obiettivo. Questo suggerisce che un movimento immaginato, come afferrare o disegnare un oggetto, stimoli il sistema nervoso in modo più profondo rispetto al gesto eseguito puramente meccanicamente, contribuendo allo sviluppo della propriocezione di quell'area corporea. Questo principio è di fondamentale importanza in ambito rieducativo.
Classificazione dei Recettori Nervosi Cutanei
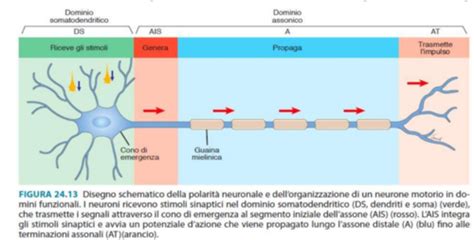
I recettori nervosi sensoriali sono formazioni distribuite in tutto il corpo con il compito di ricevere stimoli esterni e interni, trasducendoli in impulsi elettrici da inviare al sistema nervoso centrale attraverso le fibre nervose afferenti. Esistono diverse classificazioni di questi recettori. Alcuni formano strutture complesse che possono essere considerate veri e propri organi di senso.
Tra i neurorecettori sensoriali, gli esterocettori cutanei e i propriocettori rivestono un'importanza particolare dal punto di vista posturologico. Essi sono sensibili a stimoli esterni come il tatto, la pressione cutanea e altre variazioni meccaniche, anche se di derivazione interna. Possiedono diverse caratteristiche strutturali e di adattamento, e possono indurre variazioni organiche locali e, in alcuni casi, generali.
Recettori Interstiziali
Questi recettori, per lo più terminazioni nervose libere e non capsulate, sono diffusi ovunque nella cute, nei tessuti connettivi e negli interstizi muscolari. Rappresentano la famiglia di recettori nervosi di gran lunga più numerosa. Nell'innervazione muscolare umana, solo circa il 25% delle fibre sensitive deriva dai recettori noti (Golgi, Ruffini, Pacini, Paciniformi - fibre tipo I e II), mentre la restante parte origina dai "recettori interstiziali" (fibre tipo III e IV). Circa il 90% di questi ultimi sono amielinici (tipo IV), mentre i restanti possiedono una sottile guaina mielinica (tipo III).
Agiscono più lentamente rispetto ai recettori di tipo I e II. In passato, sono stati considerati prevalentemente nocicettori, termocettori e chemocettori, sensibili a vibrazioni di bassa frequenza. Tuttavia, la ricerca attuale indica che molti di essi sono multimodali e, in maggioranza, meccanocettori, sensibili a tatto, pressione e trazione. Possono essere ulteriormente suddivisi in due sottogruppi in base alla loro soglia di attivazione tramite stimoli pressori: quelli a bassa soglia (Low-Threshold Pressure - LTP) e quelli ad alta soglia (High-Threshold Pressure - HTP).
L'attivazione, in specifici stati patologici, di recettori interstiziali sensibili sia a stimoli dolorifici che meccanici (in maggioranza HTP) può generare sindromi dolorose anche in assenza di classiche irritazioni nervose, come le compressioni radicolari. Questo network sensoriale, oltre alla funzione di rilevamento del posizionamento e del movimento corporeo, influenza il sistema nervoso autonomo, modulando funzioni come la pressione sanguigna, il battito cardiaco e la respirazione, adattandole alle esigenze tissutali locali.
Recettori dei Follicoli Piliferi
Questi esterocettori cutanei sono costituiti da fibre nervose sensitive non capsulate che si avvolgono a spirale attorno alla radice del pelo cutaneo. Sono particolarmente sensibili al movimento del pelo, con un rapido adattamento.
Cellule o Dischi di Merkel (Recettori SAI)
Questi esterocettori cutanei sono formati da grosse cellule, ricche di mitocondri, con una terminazione espansa e non capsulata connessa a una fibra nervosa mielinica di grande diametro (tipo A beta - tipo II). Presentano un lento adattamento e una bassa soglia di attivazione. Si trovano in corrispondenza delle creste epidermiche, al di sotto dello strato basale, e sono spesso raggruppati a formare il recettore a cupola di Iggo, estremamente sensibile.
Le cellule di Merkel, insieme ai corpuscoli di Meissner, svolgono un ruolo importante nella determinazione delle caratteristiche spaziali delle sensazioni tattili. Essendo sensori proporzionali (sensori P), trasformano un aumento della pressione cutanea in un numero proporzionalmente maggiore di potenziali d'azione al secondo. La trasmissione degli impulsi avviene tramite una singola fibra nervosa; pertanto, solo l'eccitazione contemporanea di più cellule di Merkel adiacenti viene trasmessa come segnale al midollo spinale (convergenza di segnali).
Corpuscoli Ramificati e Lanceolati di Meissner (Recettori RA)
Questi esterocettori cutanei, situati tra epidermide e derma, sono particolarmente sensibili al tatto. Sono costituiti da una terminazione nervosa capsulata (da tessuto connettivo) e allungata, associata a una tortuosa fibra nervosa mielinica di grande diametro (tipo A beta - tipo II). Presentano un rapido adattamento.
🖐🏾🖐🏻 LA PELLE - Epidermide, Derma e Ipoderma: Funzioni e Strutture (Scienza, Il Corpo Umano)
Ciò significa che dopo un breve periodo, non reagiscono a una pressione persistente, ma solo alle sue variazioni. Quanto più rapida è la variazione di pressione, tanto più intensamente la fibra nervosa scarica (sensori differenziali o sensori D). I corpuscoli di Meissner sono molto numerosi nelle regioni cutanee glabre (prive di peli), come i polpastrelli, le labbra e i piedi.
Corpuscoli Ramificati di Ruffini (Recettori SAII)
Questi recettori sono costituiti da terminazioni nervose capsulate e pluriramificate, dove termina una fibra nervosa mielinica di grande diametro (tipo A beta - tipo II) e numerose fibre connettivali. Si trovano negli strati cutanei e tissutali profondi, nelle capsule articolari (strati superficiali), nei legamenti e nella dura madre.
I corpuscoli di Ruffini agiscono sia come esterocettori cutanei che come propriocettori. Reagiscono maggiormente agli stimoli veloci e presentano un lento adattamento statico (sensori PD proporzionali-differenziali), rendendoli importanti nel segnalare stimoli tattili e pressori prolungati e intensi. Sono particolarmente sensibili alle forze tangenziali. Come recettori articolari (meccanocettori di stiramento), segnalano la direzione e la velocità del movimento, nonché la posizione dell'articolazione, insieme ai meccanorecettori a rapido adattamento sensibili all'accelerazione.
Corpuscoli Lamellari di Vater-Pacini e Paciniformi
Questi sono corpuscoli di grandi dimensioni (fino a 4 mm di lunghezza), la cui capsula presenta numerosi strati di cellule appiattite che circondano una fibra nervosa mielinica di grande diametro (tipo A beta - tipo II). La fibra nervosa perde la guaina mielinica una volta entrata nel corpuscolo.
Si trovano immediatamente al di sotto della cute, nel tessuto adiposo ipodermico, nei tessuti connettivi profondi (generalmente più profondi dei corpuscoli di Ruffini), nelle capsule articolari (strati profondi), nei legamenti spinali e nelle giunzioni muscolo-tendinee. Sono esterocettori cutanei e propriocettori.
Organi Muscolo-Tendinei del Golgi
Questi propriocettori sono costituiti da una capsula connettivale allungata (1 mm) contenente numerose terminazioni nervose finemente arborizzate di fibre nervose ad alta velocità di conduzione (gruppo Ib). Sono situati nel tessuto connettivo denso di legamenti e capsule articolari, e nelle giunzioni miotendinee (la porzione del tendine che ospita le espansioni connettivali con cui terminano le fibre muscolari).
Gli organi del Golgi registrano la tensione muscolo-tendinea e la contrazione muscolare attiva. Se la soglia di attivazione (100-200 grammi) viene superata, innescano il riflesso spinale miotatico inverso, che provoca il rilassamento delle fibre muscolari interessate e la contrazione di quelle del muscolo antagonista. Essi proteggono le strutture tendinee, legamentose e capsulari da eccessive contrazioni muscolari.
In ambito posturologico, la loro funzione è quella di segnalare ai centri superiori la tensione media sviluppata da un gruppo selezionato di unità motorie (meccanocettori ad adattamento statico o tonico), fungendo da meccanismo di controllo riflesso della forza di contrazione muscolare (feed-back) e di programmazione della stessa da parte dei centri motori (feed-forward). Poiché gli organi del Golgi hanno una soglia di attivazione notevolmente superiore a quella dei fusi neuromuscolari, la loro azione è meno immediata e frequente.
Fusi Neuromuscolari
Questi recettori sensoriali sono diffusi nella muscolatura di tutto l'organismo, con una maggiore concentrazione nelle vicinanze delle giunzioni muscolo-tendinee dei muscoli posturali, ma anche nei muscoli dell'occhio, della mano e nei muscoli elevatori della mandibola.
Sono strutture formate da una capsula connettivale affusolata (3-10 mm di lunghezza) contenente 3-12 (fino a 36 nei muscoli masticatori) fibre muscolari specializzate (fibre intrafusali) poste parallelamente alle normali fibre muscolari (extrafusali). Le fibre intrafusali sono connesse alle estremità all'endomisio delle fibre extrafusali o al tendine.
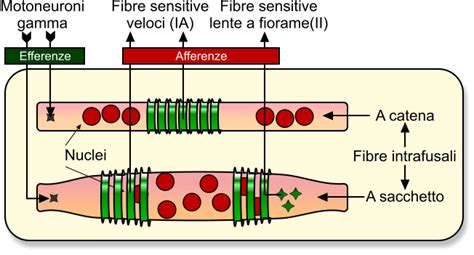
Il fuso neuromuscolare è ricco di vasi sanguigni e nervi; più della metà delle fibre di un nervo muscolare riguarda i fusi neuromuscolari. Esistono due tipi principali di fibre intrafusali: a sacco (3-5 per fuso, con circa 100 nuclei nel rigonfiamento centrale) e a catena (8-10 per fuso, con nuclei disposti a catena nella porzione centrale).
La parte recettrice del fuso si trova nella zona centrale, dove terminazioni nervose di nervi sensitivi si intrecciano con un liquido gelatinoso. Le fibre sensitive dei fusi neuromuscolari sono di due tipi: tipo Ia (primarie, trasmissione più veloce dell'organismo, una per fuso, avvolte a spirale al centro) e tipo II (secondarie, più lente e di diametro inferiore, situate più perifericamente e prevalentemente nelle fibre a catena).
Queste fibre sensitive sono eccitate da minimi allungamenti (soglia di attivazione circa 3 grammi) della zona centrale del fuso o dell'intero muscolo, e per contrazioni delle porzioni polari delle fibre intrafusali tramite fibre motorie gamma. Le terminazioni primarie (Ia) rispondono in modo dinamico (adattamento dinamico o fasico), segnalando la velocità dello stiramento. Le terminazioni secondarie (II) reagiscono in modo statico (adattamento statico o tonico), trasmettendo segnali per tutta la durata dell'eccitazione e informando sull'entità dello stiramento.
Alle estremità del fuso si trovano placche motrici formate da fibre nervose motorie mieliniche, più sottili delle fibre muscolari, derivanti dai motoneuroni gamma delle corna anteriori del midollo spinale. Queste fibre gamma si attivano per modulare la tensione delle fibre intrafusali, influenzando la sensibilità del fuso.